
5.13: Natural Selection
- Page ID
- 12114
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)
What is fitness?
Does this type of fitness have anything to do with natural selection? Usually not. There are countless ways in which an organism can be more "fit," or better adapted to its habitat. And we probably do not know about most of these adaptations.
Natural Selection
Natural selection occurs when there are differences in fitness among members of a population. As a result, some individuals pass more genes to the next generation. This causes allele frequencies to change.
Sickle Cell and Natural Selection
The example of sickle-cell anemia is described in the Figure below and Table below. It shows how natural selection can keep a harmful allele in a gene pool.
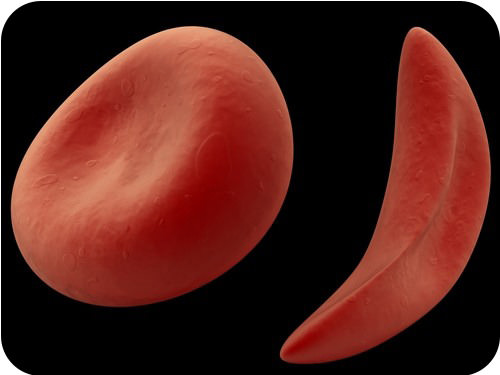 Sickle Cell and Natural Selection. Notice the normal-shaped red blood cell on the left, and the sickle-shaped cell on the right.
Sickle Cell and Natural Selection. Notice the normal-shaped red blood cell on the left, and the sickle-shaped cell on the right.| Genotype | Phenotype | Fitness |
|---|---|---|
| AA | 100% normal hemoglobin | Somewhat reduced fitness because of no resistance to malaria |
| AS | Enough normal hemoglobin to prevent sickle-cell anemia | Highest fitness because of resistance to malaria |
| SS | 100% abnormal hemoglobin, causing sickle-cell anemia | Greatly reduced fitness because of sickle-cell anemia |
Here’s how natural selection can keep a harmful allele in a gene pool:
- The allele (S) for sickle-cell anemia is a harmful autosomal recessive. It is caused by a mutation in the normal allele (A) for hemoglobin (a protein on red blood cells).
- Malaria is a deadly tropical disease. It is common in Africa south of the Sahara, South and Southeast Asia and Northern Brazil.
- Heterozygotes (AS) with the sickle-cell allele are resistant to malaria. Therefore, they are more likely to survive and reproduce. This keeps the S allele in the gene pool.
- There are three alleles found in Africa. A fourth thought to have arisen independently is found in India and Saudi Arabia.
- The allele (S) for sickle-cell anemia found in African descended people in the Western Hemisphere is believed to have come from West Africa as a result of the slave trade.
The sickle-cell example shows that fitness depends on phenotypes. It also shows that fitness may depend on the environment. What do you think might happen if malaria was eliminated in a population with a relatively high frequency of the S allele? How might the fitness of the different genotypes change? How might this affect the frequency of the S allele?
Natural Selection and Polygenic Traits
Sickle-cell trait is controlled by a single gene. Natural selection for polygenic traits is more complex, unless you just look at phenotypes. Three ways that natural selection can affect phenotypes are shown in Figure below.
- Stabilizing selection occurs when phenotypes at both extremes of the phenotypic distribution are selected against. This narrows the range of variation. An example is human birth weight. Babies that are very large or very small at birth are less likely to survive. This keeps birth weight within a relatively narrow range.
- Directional selection occurs when one of two extreme phenotypes is selected for. This shifts the distribution toward that extreme. This is the type of natural selection that the Grants observed in the beak size of Galápagos finches.
- Disruptive selection occurs when phenotypes in the middle of the range are selected against. This results in two overlapping phenotypes, one at each end of the distribution. An example is sexual dimorphism. This refers to differences between the phenotypes of males and females of the same species. In humans, for example, males and females have different heights and body shapes.
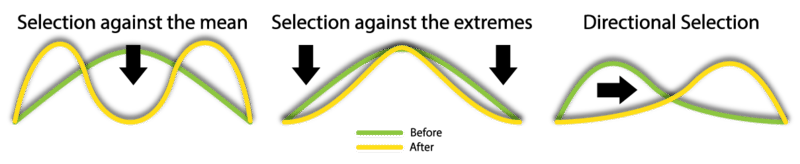 Natural selection may affect the distribution of a polygenic trait. These graphs show three ways this can happen.
Natural selection may affect the distribution of a polygenic trait. These graphs show three ways this can happen.Summary
- Natural selection occurs when there are differences in fitness among members of a population.
- Natural selection for a polygenic trait changes the distribution of phenotypes. It may have a stabilizing, directional, or disruptive effect on the phenotype distribution.
Review
- What is natural selection and what are its effects on allele frequencies?
- Describe three types of natural selection for a polygenic trait.
- How does the recessive sickle-cell allele stay in the gene pool?
On the distribution of the allele for sickle cell material has been added by LibreTexts (CC:BY) based on
Chebloune Y, Pagnier J, Trabuchet G, Faure C, Verdier G, Labie D, et al. Structural analysis of the 5’ flanking region of the beta-globin gene in African sickle cell anemia patients: further evidence for three origins of the sickle cell mutation in Africa. Proc Natl Acad Sci USA. 1988;85:4431–5.
Serjeant GR, Ghosh K, Patel J. Sickle cell disease in India: A perspective. Indian J Med Res. 2016 Jan;143(1):21-4.
Kulozik AE, Wainscoat JS, Serjeant GR, Kar BC, Al-Awamy B, Essan GJ, Falusi AG, Haque SK, Hilali AM, Kate S, et al. Geographical survey of beta S-globin gene haplotypes: evidence for an independent Asian origin of the sickle-cell mutation. Am J Hum Genet. 1986 Aug;39(2):239-44.
| Image | Reference | Attributions |
|
[Figure 1] | Credit: Zachary Wilson Source: CK-12 Foundation License: CC BY-NC 3.0 |
 |
[Figure 2] | Credit: Image copyright Sebastian Kaulitzki, 2013;U.S. Fish and Wildlife Service Source: http://www.shutterstock.com ; commons.wikimedia.org/wiki/File:Arctic_Hare.jpg License: Used under license from Shutterstock.com;Public Domain |
|
[Figure 3] | Credit: Zachary Wilson Source: CK-12 Foundation License: CC BY-NC 3.0 |